

From Histones to a Symphony in 16 Bytes
How biological complexity and assembly code exploit the chaos of their environments.

The Crisis of Reductionism
In How Life Works: A User’s Guide to the New Biology, former Nature editor Philip Ball describes his early frustration with biological literature. Researchers might isolate a single gene or map out the crystal structure of a receptor. However, these papers almost always end with a hidden admission of ignorance: in isolation, it is profoundly difficult to discern what these findings actually mean for a living organism.
The traditional sequence-to-structure-to-function simplification has driven decades of progress, but it remains fundamentally incomplete. By treating biological systems as linear, deterministic machines, we often miss the underlying rules of how life actually computes.
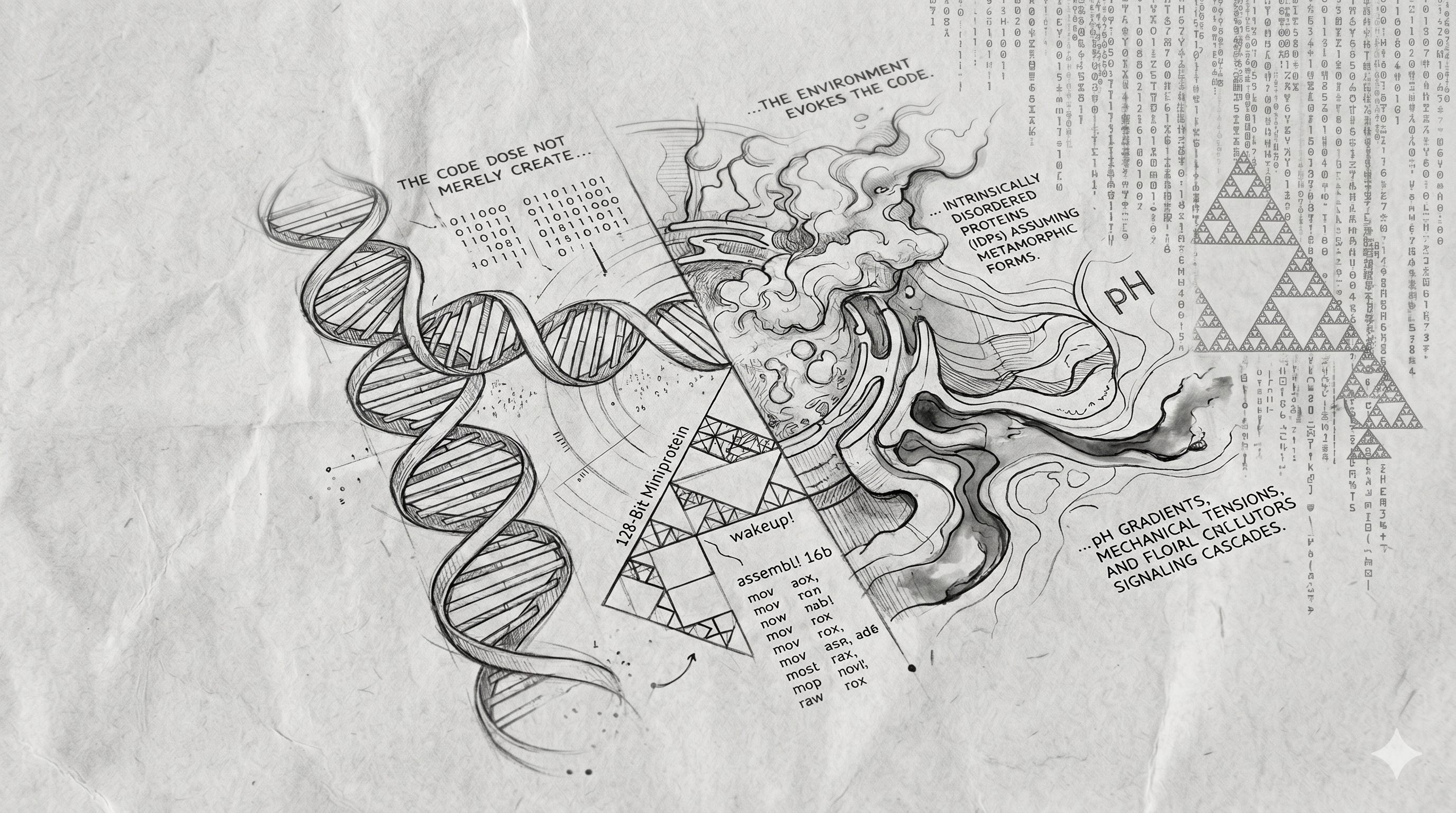
Simple, But Not Simpler
In higher education, the genome is often taught as a pristine, unidirectional blueprint. Students are introduced to the Central Dogma, a paradigm that reduces biology to a clean, feed-forward factory floor where DNA is transcribed to RNA, translated to protein, and folded into a predictable geometry to execute a singular task.
But the reality of transcription is contextual. Because the genome is a dense, tangled polymer, a single gene cannot simply switch on; the surrounding architecture of the cell must undergo an extensive physical remodelling. To expose a chromosomal sequence, DNA must be unwrapped from its spools.
Ultimately, genetic accessibility is a dialogue between the cell’s physical microenvironment and its internal machinery. While chromatin architecture, epigenetic marks, and transcription factors establish the core code, localised pH gradients, mechanical tensions, and signalling cascades dictate the final context of how that code is read.
Shifting Clouds
The limits of pure reductionism become glaringly obvious in the study of Intrinsically Disordered Proteins (IDPs). These molecules entirely lack a fixed, native three-dimensional structure. They exist within the crowded intracellular environment as fluid, shifting ensembles, defying the classic lock-and-key model.
This does not invalidate traditional sequence-to-structure biology; structure-function relationships remain highly predictive for the vast majority of classic enzymes. Rather, IDPs reveal the boundaries of that paradigm. They show that structure can be a fluid variable rather than a fixed constant, acting as molecular sensors that shape-shift in response to cellular signals.
Amplifying Minor Deltas
An amino acid sequence is not a set of explicit behavioural instructions; it is a highly compressed, latent representation of potential energy states.
Its actual function is amplified, modulated, and ultimately defined by its continuous interaction with the world around it. A minor mutation in an enzyme’s sequence might only slightly tilt its static shape. However, the cellular environment can take this tiny shift and amplify it drastically. This environment might hyper-charge the enzyme’s catalytic power, or, conversely, instantly mark it for destruction by the proteasome. Information does not reside solely in the sequence, but in the interaction between code and substrate.
The Digital Cytoplasm
To observe this biological truth mirrored in artificial architecture, one must glance backward into the history of computing: specifically, to the raw, uninsulated landscape of real-mode DOS.
At the Outline Demoparty in May of 2026, the digital artist HellMood released wakeup! 16b - a masterwork of algorithmic density written in exactly sixteen bytes of x86 assembly language.
To most software engineers, the assertion that sixteen bytes could manifest anything meaningful is absurd. To put this in perspective, sixteen bytes is smaller than a single cryptographic key, or the length of “the quick brown “. When executed within the primitive playground of vintage hardware, those sixteen bytes unfurl into an infinite, self-healing cascade of algorithmic green rain while simultaneously synthesising a rhythmic, driving techno soundtrack.
How does a piece of code so minuscule achieve an expression so vast? Like a cell exploiting chemistry, the programme exploits the physics of the machine around it.
As HellMood details in his technical writeup, the code completely ignores bounds-checking, memory allocation, and explicit control flow. Instead, it instructs the processor to step backward through memory. It treats the uniform BIOS screen lattice as a clean geometric baseline for its visual Sierpinski-like fractals, while weaponising raw machine noise to supply the data needed to generate its gritty, industrial audio.

Figure 1. The Matrix Rain from HellMood’s audio-video 16 byte masterpiece.
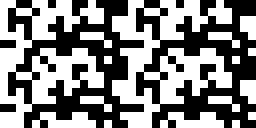
Figure 2. Visual representation of the exact 128 zeroes and ones in the programme.
The Medium and the Message
If you extract those sixteen bytes of assembly and attempt to execute them within the sterile vacuum of a modern 64-bit operating system, they denature instantly. They are meaningless without the exact environment in which they evolved.
This is how we must view gene expression and protein function. The sequence defines the landscape of possibilities, but the environment selects the specific trajectory through it.
In both biology and low-level computation, complexity is rarely written explicitly into the code itself. Instead, the code establishes elegant constraints that allow rich, unpredictable behaviour to emerge through interaction with the environment.
Evolution exploits this principle relentlessly: genomes outsource their complexity to physics, chemistry, and cellular architecture in much the same way demoscene programmers outsource behaviour to hardware quirks and memory states.
You can witness the emergent output of those 16 bytes here.
#bits #atoms